What skeletal muscle taught us about obesity, oxidative stress, and gestational diabetes.
Pregnancy is one of the most demanding metabolic challenges the body will ever face. And it’s not just the hormones. Blood volume and cardiac output increase by up to 50%. Breathing rate, lung capacity, kidney filtration (and size!), all increase dramatically. Metabolically, the development of the fetus reigns: early in pregnancy, insulin sensitivity increases to store carbohydrates and fats, while later, insulin resistance rises to ensure plenty of glucose for the rapidly growing fetus. In the last half of gestation, mom is left to rely more heavily on stored fats for energy, effectively doubling fat metabolism, a metabolic challenge in its own right.
In the face of these metabolic adaptations, many people develop complications related to metabolic disease, like hypertension (preeclampsia) or gestational diabetes. If you’ve been following along, this essay is the next in my From the Paper series, where I summarize our research studies for those outside of academia, from the beginning! Earlier essays in this series focused on my dissertation research in adults with obesity, where we studied how skeletal muscle responded to nutritional challenges like excess fats in vivo, and in cells cultured from muscle biopsies.
My next paper begins with my first project as a postdoctoral fellow at the University of Colorado Anscutz. In this study, we set out to understand how skeletal muscle responds to the metabolic challenge of pregnancy, particularly in those women with who develop gestational diabetes (PMID: 23956348).1 But what we found was that the differences were not unique to gestational diabetes at all.
I’d like to note that this research was funded by the NIH. If you’d like to see more work like this supported in the future, please take a minute to read my essays about current challenges with NIH funding and what you can do about it (here and here).
Reader’s Note: Here, I use the term “people with obesity” to reflect that obesity as a complex, chronic disease. This study defined obesity based on pre-pregnancy or first trimester BMI. The findings discussed describe group‑level patterns, not individual predictions, as does the clinical designation of obesity by BMI. People with obesity are metabolically diverse, and my ongoing research focuses on understanding this heterogeneity and its origins.
Background
This study focuses on gestational diabetes, defined as diabetes first diagnosed during pregnancy. In broad terms, this refers to difficulty regulating blood sugar (glucose), which can arise in several ways. In some cases, the pancreas doesn’t produce enough insulin. More commonly in the context of obesity, the body becomes less responsive to insulin, referred to as insulin resistance. During pregnancy, everyone becomes more insulin resistant, especially in the second half of gestation. This shift is driven by hormones produced by the placenta and helps prioritize energy for the developing fetus. However, in people with underlying risk factors, this normal shift can be amplified, progressing into gestational diabetes.
After delivery, blood glucose levels often return to normal within days or even hours. However, this does not mean the risk is over. A third of those with gestational diabetes will go on to develop type 2 diabetes within 15 years (PMID: 32404325, PMID: 33333204).2,3 One hypothesis is that pregnancy acts as a metabolic ‘stress test,’ unmasking underlying susceptibility for metabolic disease. This may help explain why pre-pregnancy obesity, affecting ~30% of individuals in the U.S., is associated with a three-fold higher risk of developing gestational diabetes.
Skeletal muscle plays a central role in this process. It accounts for a large portion of our energy expenditure and is responsible for most glucose uptake, especially in response to insulin. With gestational diabetes, skeletal muscle insulin resistance is higher than in normal glucose tolerant pregnancies (non-diabetic, PMID: 10480612).4 Outside of pregnancy, insulin resistance has been linked to altered mitochondrial metabolism and increased oxidative stress (see box, below). However, it is not known whether these same processes underlie gestational diabetes, or are evident more broadly across metabolic conditions within pregnancy.
What is oxidative stress?
Oxidative stress refers to an imbalance between the production of reactive oxygen species (oxidants) and antioxidant defenses. Reactive oxygen species (ROS) are produced naturally in our cells as a result of everyday chemical reactions, most notably when oxygen is used to generate energy during mitochondrial metabolism in the electron transport system. Other cellular processes, such as protein folding or signaling can also produce ROS, and external factors like smoking, UV radiation, or certain chemicals can increase their production.
Antioxidants act as neutralizing agents. Their job, often described as scavenging, is to keep ROS in balance. Small amounts of ROS are beneficial, carrying out important cellular functions. But if ROS accumulate faster than they can be neutralized, the balance shifts, called oxidative stress. Excess ROS can damage cellular components such as cell membranes (through lipid peroxidation, where free radicals can oxidize membrane lipids). ROS can also damage proteins involved in insulin signaling, enzymes like aconitase or mitochondrial electron transport Complex II. ROS can even damage DNA, which contributes to aging and cancer. When it comes to ROS, balance is key.
In this study, we focused on several parts of this system: ROS‑sensitive enzymes, such as aconitase, mitochondrial enzymes, and antioxidant defenses, including MnSOD and glutathione. Together, these provide a snapshot of whether cells are maintaining balance or shifting toward oxidative stress.
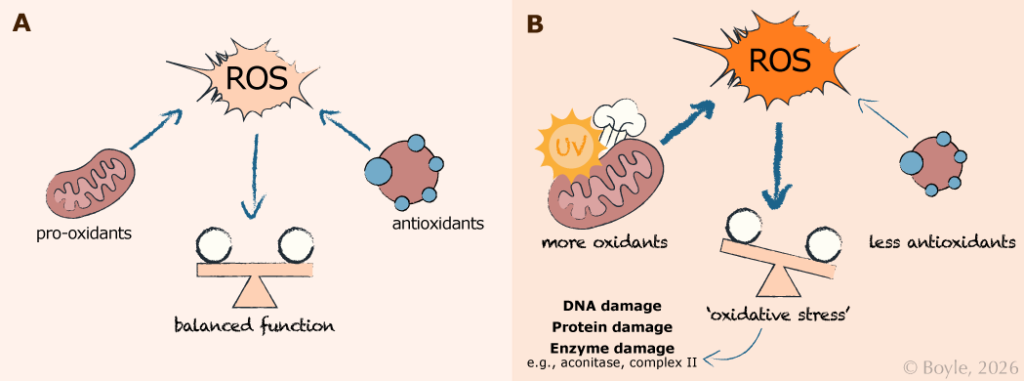
Oxidative stress arises from an imbalance between reactive oxygen species (ROS) production and antioxidant defenses. Panel A shows a balanced state. Under normal conditions, ROS are generated as a natural byproduct of cellular metabolism in the mitochondria. Typically, these ROS are effectively neutralized by antioxidants, maintaining balance and normal cellular function. Panel B shows oxidative stress. When ROS production increases, due to more mitochondrial metabolism or environmental stressors such as UV exposure or smoking, or when antioxidant capacity is reduced, this balance shifts toward oxidative stress. Excess ROS can damage DNA, proteins, and enzymes, impairing cellular and metabolic function.
The question:
Most prior work in gestational diabetes has focused on blood sugar levels and how the body responds to insulin. Whether insulin resistance in muscle from people with gestational diabetes was related to differences in mitochondrial metabolism or oxidative stress was not known. In this study, we asked:
Are the mitochondria or oxidative stress in the muscle from people with gestational diabetes different from those without diabetes?
What we did:
Because gestational diabetes often occurs alongside obesity, it can be difficult to distinguish its effects from those of obesity alone. focusing only on individuals with obesity misses potential baseline differences relative to normal‑weight pregnancy. To address this, we examined skeletal muscle samples collected during C‑section from three groups: normal‑weight with normal glucose regulation, obese with normal glucose regulation, and obese with gestational diabetes.
Mitochondrial measures
Instead of measuring mitochondrial respiration, as I described in this paper, here we assessed the maximum capacity of mitochondrial enzymes. While respiration provides a more direct measure of metabolism in living mitochondria, this enzyme approach allows analysis of frozen samples. In addition to activity of enzymes in the electron transport system, we measured markers of mitochondrial content to distinguish differences in amount of mitochondria from differences in their function.
Oxidative stress measures
We assessed oxidative stress using several complementary measures. First, we measured activity of aconitase, an enzyme that is highly sensitive to oxidative damage. We also measured levels of oxidized and reduced glutathione, a key cellular antioxidant system whose balance reflects overall oxidative stress. Finally, we measured activity of the antioxidant enzyme MnSOD (manganese superoxide dismutase).
Enzyme regulators
Lastly, we examined upstream regulators of enzyme activity, including sirtuin 3 (Sirt3), a mitochondrial enzyme that modifies other proteins. In this context, Sirt3 can increase the activity of enzymes of the electron transport system and MnSOD by removing acetyl groups.
A deeper dive into oxidant production & neutralization
Reactive oxygen species (ROS) are highly reactive molecules that include superoxide (O₂⁻) and hydrogen peroxide (H₂O₂). They can be generated at several sites within mitochondria when electrons ‘leak’ from the electron transport system and react with oxygen. For example, superoxide forms when an oxygen molecule (O₂) gains a single electron, creating a highly reactive, negatively charged oxygen molecule known as a free radical. If you’d like to learn more about reactive oxygen species and mitochondrial metabolism, this review provides an in-depth overview (PMID: 28842493).5
Antioxidant systems can neutralize ROS before they can cause damage. For example, the enzyme manganese superoxide dismutase (MnSOD) converts superoxide into oxygen (O₂) and hydrogen peroxide (H₂O₂), which is less reactive. In turn, glutathione then converts hydrogen peroxide into harmless water (H₂O). During this reaction,
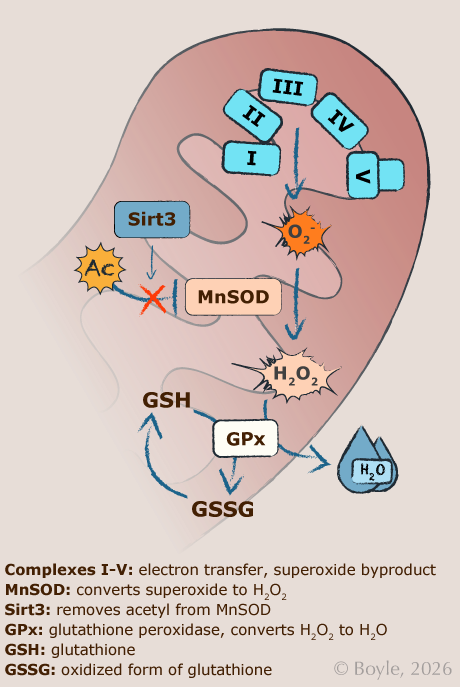
glutathione itself becomes oxidized. The balance between its reduced (GSH) and oxidized forms (GSSG), which we measured in this study, is widely used as an indicator of oxidative stress within cells.
Mitochondrial enzymes are both a source and a target of ROS. For example, Complex II can contribute to ROS production but is also vulnerable to oxidative damage, which reduces its efficiency. Other enzymes, like aconitase, are particularly sensitive to oxidative damage, resulting in reduced activity. Over time, this can create a feedback cycle: increased ROS causes more mitochondrial damage, which further increases ROS production. This oxidative stress can also trigger inflammation and is thought to contribute to the chronic, low-grade inflammation commonly observed in obesity and insulin resistance (PMID: 21686173).6
We also examined Sirt3, a mitochondrial enzyme that helps maintain this balance. Sirt3 removes small chemical tags called acetyl groups from proteins, including MnSOD and several components of the electron transport system. Removing these acetyl groups increases the activity of these proteins, strengthening antioxidant defenses and supporting efficient mitochondrial function. When Sirt3 activity is reduced, these proteins remain more highly acetylated and less active, allowing oxidative stress to build.
Figure shows mitochondrial mechanisms of reactive oxygen species (ROS) production and antioxidant defense. Electron leak from the mitochondrial electron transport system (Complexes I-V) generates superoxide (O₂⁻), which is converted to hydrogen peroxide (H₂O₂) by manganese superoxide dismutase (MnSOD). Hydrogen peroxide is then neutralized by glutathione-dependent antioxidant systems, with the GSH ratio serving as an indicator of cellular redox balance. Sirtuin 3 (Sirt3) enhances antioxidant defense and mitochondrial function by deacetylating MnSOD and electron transport proteins. Together, these pathways regulate the balance between normal mitochondrial function and oxidative stress.
What we found:
Mitochondrial measures
We found that maximal capacity of one electron transport enzyme (Complex II) was lower in both both groups with obesity (those with obesity alone, and who also had gestational diabetes) compared with the normal weight, normal glucose‑tolerant group. There were no differences in markers of mitochondrial content, indicating that the changes reflect reduced enzyme function rather than differences in mitochondrial quantity.
Oxidative stress measures
A similar pattern was observed for oxidative stress. Both groups with obesity showed evidence of greater oxidative stress compared with the normal weight group: aconitase activity was lower, and glutathione was more oxidized. MnSOD activity was also reduced, indicating lower antioxidant defenses in the groups with obesity.
Enzyme regulators
Sirt3 enzyme activity was also lower in both groups with obesity compared with the normal weight group. Sirt3 mRNA content was also lower with obesity, suggesting lower overall Sirt3 levels. When we examined acetylation levels of Sirt3 target proteins (electron transport Complex II and MnSOD), we found no differences in the Complex II, but MnSOD showed higher acetylation in both groups with obesity. This is consistent with lower enzyme activity and Sirt3 function
What it means:
In skeletal muscle during pregnancy, those with obesity (with and without gestational diabetes) showed remarkably similar patterns of reduced mitochondrial capacity and greater oxidative stress compared with the normal weight, normal glucose tolerance group. Reduced antioxidant defenses with obesity may be mediated by lower Sirt3 activity, leading to increased oxidative stress, independent of gestational diabetes.
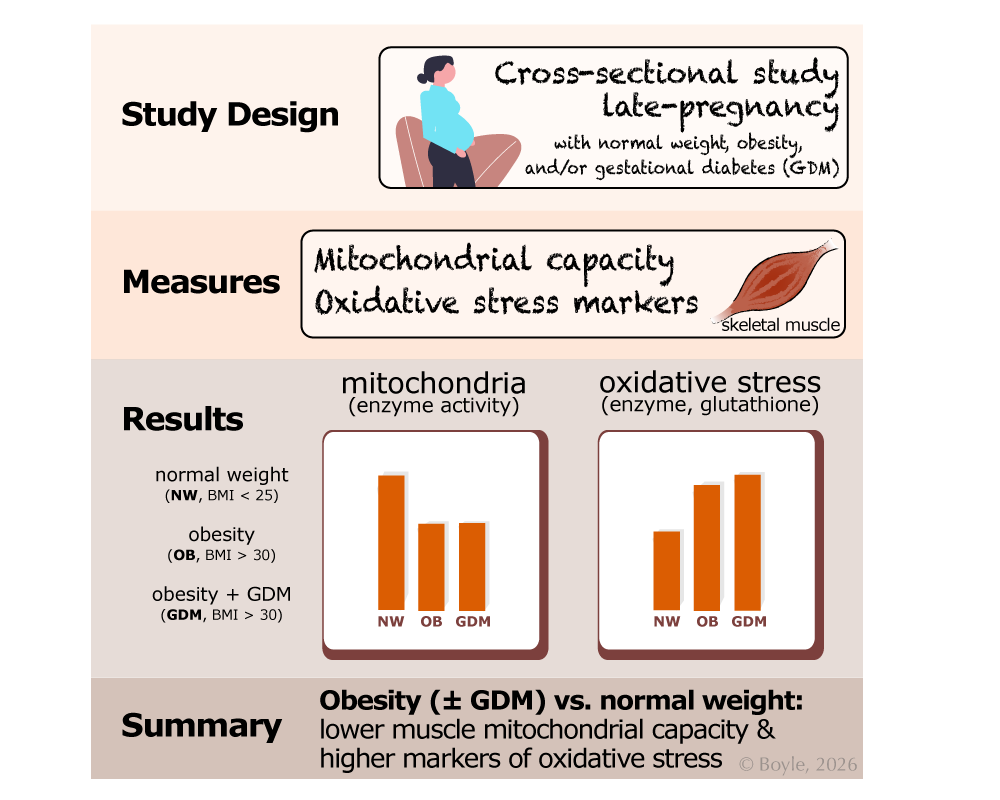
Study Overview: This was a cross-sectional study that compared skeletal muscle from people in late pregnancy with normal weight (NW), obesity (OB), or obesity with gestational diabetes (GDM). Mitochondrial capacity and markers of oxidative stress were measured. Compared with normal weight, obesity with or without GDM was associated with lower mitochondrial capacity and higher oxidative stress, with little additional effect of gestational diabetes beyond obesity.
What we still don’t know (a lot!):
This is a cross-sectional study, with samples taken at delivery. We do not know if the metabolic and oxidative stress differences we observed were present prior to pregnancy or if they developed over the course of gestation. While we do know from prior research that many people with gestational diabetes return to normal glucose tolerance after delivery, we do not know if their muscle metabolic and oxidative stress features will resolve as well.
Oxidative stress is a catch-all term that can implies greater oxidant burden, which relies on the balance of pro- and anti-oxidant mechanisms. While we measured several markers of oxidative stress, these are not a comprehensive assessment of all potential sources of pro- or anti-oxidants.
If we consider the metabolic changes that occur during pregnancy, specifically in late gestation when maternal metabolism preserves carbohydrates for the fetus and shifts maternal energy sources toward stored fats, those with obesity may face further metabolic burden. As discussed in this research summary, fats are primarily oxidized in the mitochondria. If the capacity to do so is limited, these metabolic transitions may be quite challenging. Such reduced ability to oxidize available fats may lead to heavy competition for glucose between the mother and fetus. To my knowledge, this has not been investigated and certainly warrants further study.
Overall, these data suggest that, in pregnancy, skeletal muscle insulin resistance may be dissociated from mitochondrial dysfunction and oxidative stress, which are often linked in non-pregnant individuals. And even though reduced mitochondrial capacity and oxidative stress co-existed in the groups with obesity, we do not know if oxidative stress contributed to metabolic differences or vice versa. The causal links between mitochondrial health, oxidative stress, and insulin resistance both within and outside of pregnancy, are still unclear and warrant further study.

Kristen Boyle, PhD
Dr. Kristen Boyle is Associate Professor of Pediatrics at the University of Colorado School of Medicine, where she leads the Molecular Health Lab. Her work examines how early life shapes lifelong health. She writes about how science works, how we interpret evidence, and how we can make sense of uncertainty.
Linked References
- Boyle KE, Newsom SA, Janssen RC, Lappas M, Friedman JE. Skeletal muscle MnSOD, mitochondrial complex II, and SIRT3 enzyme activities are decreased in maternal obesity during human pregnancy and gestational diabetes mellitus. J Clin Endocrinol Metab. 2013 Oct;98(10):E1601-9. doi: 10.1210/jc.2013-1943. PMID: 23956348. https://pubmed.ncbi.nlm.nih.gov/23956348/
- Vounzoulaki E, Khunti K, Abner SC, Tan BK, Davies MJ, Gillies CL. Progression to type 2 diabetes in women with a known history of gestational diabetes: systematic review and meta-analysis. BMJ. 2020 May 13;369:m1361. doi: 10.1136/bmj.m1361. PMID: 32404325. https://pubmed.ncbi.nlm.nih.gov/32404325/
- Dennison RA, Chen ES, Green ME, Legard C, Kotecha D, Farmer G, Sharp SJ, Ward RJ, Usher-Smith JA, Griffin SJ. The absolute and relative risk of type 2 diabetes after gestational diabetes: A systematic review and meta-analysis of 129 studies. Diabetes Res Clin Pract. 2021 Jan;171:108625. doi: 10.1016/j.diabres.2020.108625. Epub 2020 Dec 15. PMID: 33333204. https://pubmed.ncbi.nlm.nih.gov/33333204/
- Friedman JE, Ishizuka T, Shao J, Huston L, Highman T, Catalano P. Impaired glucose transport and insulin receptor tyrosine phosphorylation in skeletal muscle from obese women with gestational diabetes. Diabetes. 1999 Sep;48(9):1807-14. doi: 10.2337/diabetes.48.9.1807. PMID: 10480612. https://pubmed.ncbi.nlm.nih.gov/10480612/
- Wong HS, Dighe PA, Mezera V, Monternier PA, Brand MD. Production of superoxide and hydrogen peroxide from specific mitochondrial sites under different bioenergetic conditions. J Biol Chem. 2017 Oct 13;292(41):16804-16809. doi: 10.1074/jbc.R117.789271. Epub 2017 Aug 24. PMID: 28842493. https://pubmed.ncbi.nlm.nih.gov/28842493/
- Fernández-Sánchez A, Madrigal-Santillán E, Bautista M, Esquivel-Soto J, Morales-González A, Esquivel-Chirino C, Durante-Montiel I, Sánchez-Rivera G, Valadez-Vega C, Morales-González JA. Inflammation, oxidative stress, and obesity. Int J Mol Sci. 2011;12(5):3117-32. doi: 10.3390/ijms12053117. Epub 2011 May 13. PMID: 21686173. https://pubmed.ncbi.nlm.nih.gov/21686173/